
|
|
Cannabinoids and Memory
Cannabis sativa and humans have a long history together, dating back several thousand years when the plant was first discovered and used by Indian and Chinese civilizations for recreational, medicinal, religious, and industrial purposes. In modern times, it has been banned from recreational and medicinal use in many countries, and the issue of its safety and value has been a hot source of controversy. This at first deterred research into the effects of the plant and its active ingredient, delta-9-tetrahydrocannabinol (THC), on the human body. More recently, however, the discovery of receptors in animals and humans that bind THC and other cannabinoids, some of which are endogenous, has led to exciting new knowledge about the physiology of animal systems. As cannabinoid receptors are most abundant in the brain and peripheral nervous system, this discovery has most significantly impacted the field of neurobiology. Cannabinoid receptors have been found, along with endogenous agonists, in a wide variety of animal species ranging from mollusks and leeches to mammals, including humans. In addition, the receptors and agonists are structurally similar across species. This is quite extraordinary since vertebrate and invertebrate evolutionary history diverged more than 500 million years ago. In my opinion, finding cannabinoid receptors and their endogenous agonists in both suggests that they were preserved because of their importance in serving an irreplaceable and fundamental function in animal physiology and, more specifically, in animal neurophysiology. Otherwise, one would expect such divergent evolutionary trends to modify or even eradicate the cannabinoid system. These receptors are abundant in the human brain, and they are especially plentiful in the hippocampus, a region of the brain responsible for some types of learning and memory. This is not surprising, since a well-known side effect of smoking or ingesting cannabis is the disruption of short term and working memory, both of which are mediated by the hippocampus. The plethora of recent research on cannabinoids has shown that the endocannabinoid system in neurons is an important component in the regulation and operation of learning and memory. Understanding the role of these receptors and their agonists in neural signaling may uncover many of the secrets of how animals learn. We first look at general studies on the effects of cannabinoids on animal learning and memory. We then examine the specific cellular phenomena associated with the endocannabinoid system that may correlate with these observed effects. The Endocannabinoid SystemThe first breakthrough into cannabinoid research was the identification of the two cannabinoid receptors in the body named CB1 and CB2 that THC binds to, producing the observed effects of cannabis use. This launched a search for agonists other than THC that occur naturally in the body, and so far two endogenous cannabinoids have been identified—anandamide (arachidonyl ethanolamide) and 2-AG (2-arachidonyl glycerol). This told researchers that an endocannabinoid system exists in the body, and that the effects of THC are likely caused by a disruption of this naturally occurring system. After this realization, receptors and their agonists were found in vertebrate and invertebrate species (Benson et. al., 1999), showing researchers that this is an old system important in the physiology of many different animals at a fundamental level. It was also determined from their synthesis that anandamide and 2-AG are related to prostaglandins, which are present in the earliest of species (Di Marzo et. al., 1996). The next step was to determine the properties of the various components of the endocannabinoid system and its location(s) in the body. The CB1 and CB2 receptors differ in their structure, affinity for ligands, and distribution in the human body. CB1 is found primarily in the CNS/PNS, but can also be found along with CB2 in the heart, liver, spleen, cells of the immune system, and sperm (Benson et. al., 1999). This finding also showed that cannabinoids affect many systems in the body, but are particularly prevalent in the nervous system. Several synthetic cannabinoids have been developed that bind to CB1 with varying affinity and have been used in radioligand studies along with behavioral studies to identify cannabinoid receptors in brain regions involved in pain, learning, memory, motor control, pleasure, hunger, and higher level processing in association cortex. These same studies have found especially high numbers of receptors in the hippocampus, the region of the brain notorious for its role in learning and memory. This role in learning in memory, it is now apparent, was the most exciting new knowledge gained by understanding how the system works. Exogenous Cannabinoids Disrupt Learning and MemoryThere are several well-replicated experiments on the effects of THC on learning and memory that help to understand the function of cannabinoids in these processes. Many studies in humans have shown that cannabis intoxication is analogous to a temporary lesion in the hippocampus (Benson et. al., 1999). It impairs short-term memory and working memory, processes thought to take place in the hippocampus. There is also evidence that THC impairs spatial working memory in rats performing maze tasks, but not reference memory, which is also analogous to a reversible hippocampal lesion (Bowers et. al., 1997). This has led to an experiment to test this theory.
Hippocampal lesions can also alter the strategy that a rat uses in performing the DNMS task; they will begin to prefer one lever to another, rather than trying to remember which one is correct (Deadwyler & Hampson, 1998). This effect was also observed during a study in rats intoxicated with THC. They altered their strategy to prefer a lever, just as did rats with hippocampal lesions. We can conclude that proper operation of the endocannabinoid system is required for hippocampus-dependent memory tasks. As we will see, THC acts in a non-specific manner, disrupting the normal operation of this system. Its effect on memory, however, may not take place solely through the hippocampus. THC also affects portions of association cortex, as there are many cannabinoid receptors found there (Breivogel & Childers, 1998). Experiments on local administration of THC in prefrontal cortex show that it decreases rat performance in spatial tasks and increases dopamine and norepinephrine levels (Andrusiak et. al., 1997). Administering N-methyl-D-aspartate (NMDA) antagonist HA966 has no effect alone, but reverses both the THC-induced memory impairment and the dopamine increase, while not affecting the increased norepinephrine levels. This leads us to believe that THC causes short term memory impairment not only through actions in the hippocampus, but also by increasing dopamine levels in prefrontal cortex. This in turn suggests that, in addition to the hippocampus, the normal behavior of cannabinoids in prefrontal cortex is also necessary for some learning and memory-related tasks. Cellular MechanismsOne consistent finding in studies of the type above is that where synthetic and/or exogenous cannabinoids such as THC are used to obtain results, endogenous cannabinoids like anandamide fail to produce the same results. This cannot be explained without a better understanding of the cellular processes that take place regarding endocannabinoids. Experiments done in the last few years towards determining this cellular mechanism have shown that in any neural circuits where they are present endocannabinoids may participate in a retrograde messenger system whose goal is presynaptic inhibition. Endocannabinoids serve as the messengers in this system, and CB1 serves as the receptor that initiates the inhibition. This is especially important in signaling between neurons in the hippocampus, where strengthening and weakening of neural connections, thereby reorganizing neural circuits, is thought to be a cellular correlate of learning and memory.
The figure on the left shows results in levels of 2-AG in stimulated and unstimulated hippocampal slices. The stimulated slices had higher levels of 2-AG (a). The effect is blocked by tetrodotoxin (b), a Na+ channel blocker, and is blocked by removal of Ca2+ from solution (c), confirming that, like anandamide, 2-AG release requires both depolarization and Ca2+ influx into the postsynaptic cell. The experiment did not detect any influence in anandamide levels from stimulation of Schaeffer collaterals, but this may just indicate that anandamide is not released through this particular pathway. We can thus hypothesize that in vivo, CA1 pyramidal cells (and numerous other types of neurons throughout the brain) will release endocannabinoids in response to presynaptic stimulation, completing the first step of the retrograde messenger process. The endocannabinoids are also inactivated quickly, allowing for a localized, short-lived response. This can explain why in most studies, exogenous cannabinoids exert effects while endogenous ones do not; the endogenous cannabinoids are rapidly degraded by the body and do not have the opportunity to act. We now turn to the effects of their release.
The figure above shows an experiment testing long-term potentiation (LTP) of hippocampal CA1 pyramidal cells (Piomelli et. al., 1997). After application of a high frequency stimulus, or tetanus, the control slice potentiated. Slices bathed in 2-AG showed an initial increase in excitatory post-synaptic potentials (EPSPs), but this increase was transient, and no LTP occurred. The experiment also found that 2-AG alone has no effect on the basal EPSP, and that a CB1 receptor antagonist blocked the inhibition of LTP by 2-AG. We can therefore conclude that 2-AG inhibits LTP, and binding to CB1 is necessary for this effect. Understanding the effects of binding to a cannabinoid receptor on the presynaptic cell is the remaining task.
From studies on the mechanisms of LTP in CA1 pyramidal cells and also habituation in Aplysia, we know that reducing cAMP levels in a cell should cause decreased activity of protein kinases (such as PKA) and lead to opening of K+ channels, producing action potentials with shorter duration. With shorter action potentials, we also get less diffusion into the cell of Ca2+ and, thus, less neurotransmitter release. This theory has been tested and verified. It has been demonstrated that cannabinoids modulate K+ current in hippocampal neurons in a cAMP-dependent manner (Childers et. al, 1995). It has also been shown in purkinje cells that cannabinoids inhibit presynaptic Ca2+ influx (Kreitzer & Wade, 2001). All that remains is to show that cannabinoids act in a retrograde fashion to inhibit presynaptic cells. Excitation and Inhibition by Endocannabinoids
Depolarization-induced suppression of excitation (DSE) is the phenomenon where
depolarization of the postsynaptic cell
We have thus identified mechanisms by which endocannabinoids, acting as retrograde messengers for presynaptic inhibition, can act to both strengthen and weaken the connections between neurons. They strengthen connections by decreasing presynaptic inhibition, and weaken connections by decreasing presynaptic excitation. One type of simple learning, reflex conditioning, may be better understood through the action of endocannabinoids. The nictitating membrane reflex occurs in rabbits. It consists of pulling the nictitating membrane over the eye in response to stimulation. This reflex can be conditioned to respond to a tone by paring it with an airpuff, the unconditioned stiumulus. The current model for this requires purkinje cells to decrease firing, thus decreasing their inhibition of motor effectors, to produce the conditioned response. This is thought to happen upon the coincident activation of parallel and climbing fibers by the conditioned stimulus and the unconditioned stimulus, or the airpuff paired with the tone, but exactly how this happens has not been determined. We have seen that depolarization of purkinje cells causes DSE through endocannabinoid release. This may be the missing piece of the puzzle, as it is plausible that purkinje cells release endocannabinoids in response to the pairing of climbing and parallel fibers, inhibiting their own activation by them and producing the motor response. This application to a very simple type of learning can, perhaps, be generalized to the processes that take place in the hippocampus. A possible purpose in the hippocampus for presynaptic retrograde inhibition may be to forget “stored stimuli following retrieval…Another possibility is that cannabinoid receptor activation prevents stored information from being ‘overwritten’ by extraneous stimuli” (Deadwyler & Hampson, 1998). The inhibitory effects, expressed as decreases in glutamate release, can explain the memory suppressive effects of exogenous cannabinoid application. Although its exact role in the hippocampus is still to be determined, the endocannabinoid system shows promise of being a companion to LTP in generating the processes of learning and memory in the brain. Summary and Conclusions Experiments on cannabinoids to date have revealed that their proper operation is necessary for the neurobiological process responsible for some learning and memory. They have been shown to disrupt memory and inhibit LTP in hippocampal slices. We have also seen that they act as retrograde messengers for presynaptic inhibition in all areas of the brain, but their exact role and effect on neural circuits has not been completely elucidated. Since they are capable of both strengthening and weakening neural connections, a good hypothesis is that they engender learning and memory by reorganizing neural circuits. Donald Hebb said in his famous 1949 book, The Organization of Behavior: There are memories which are instantaneously established, and as evanescent as they are immediate… If some way can be found of supposing that a reverbatory trace might cooperate with the structural change, and carry the memory until the growth change is made, we should recognize the theoretical value of the trace which is an activity only, without having to ascribe all memory to it. This reverbatory trace that Hebb proposes is a short-lived reorganization of neural circuits to represent memory, and learning is the acquisition of these reverbatory traces. LTP has already been shown to alter the strength of connections between neurons in the hippocampus and, thus, is thought to be necessary for learning and memory. From our discussion of the endocannabinoid system, it too should participate in learning and memory, in light of its ability to reorganize neural circuits. Testing this hypothesis should provide new and exciting information about the way the brain works. References
Christie, M., Vaughan, C. Cannabinoids act backwards. Nature. 410, 527-530 (2001)
Kreitzer, A., Regehr, W. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto purkinje cells. Neuron. 29, 717-727 (2001)
Nicoll, R., Wilson, R. Endogenous cannabinoids mediate retrograde signaling at hippocampal synapses. Nature. 410, 588-592 (2001)
Marijuana and Medicine, Assessing the
Science Base. Benson, J. et. al., ed.:
Breivogel, CS., Childers, SR. The functional neuroanatomy of brain cannabinoid receptors. Neurobiology of Disease. 5(6), 417-431 (1998)
Deadwyler, S., Hampson, R. Role of cannabinoid receptors in memory storage. Neurobiology of Disease. 5(6), 474-482 (1998)
Felder, C., Glass, M. Cannabinoid receptors and their endogenous agonists. Annual Review of Pharmacology and Toxicology. 38, 179-200 (1998)
Andrusiak, E. et. al. Delta 9-tetrahydrocannabinol increases prefrontal cortical catecholaminergic utilization and impairs spatial working memory in the rat: blockade of dopaminergic effects with HA966. Neuropsychopharmacology. 16(6), 426-432 (1997)
Piomelli, D., Schweitzer, P., Stella, N. A second endogenous cannabinoid that modulates long-term potentiation. Nature. 388, 773-778 (1997)
Di Marzo, V., et. al. Potential biosynthetic connections between the two cannabimimetic eicosanoids, anandamide and 2-arachidonoyl-glycerol, in mouse neuroblastoma cells. Biochemical & Biophysical Research Communications. 277, 281-288 (1996)
Childers, S. et. al. Cannabinoids modulate voltage sensitive potassium A-current in hippocampal neurons via a cAMP dependent process. Journal of Pharmacology and Experimental Therapeutics. 273, 734-743 (1995)
Author: Krishna
|
||
Click here to refer this page to a friend and let freedom grow Home | Mission | Patient
Resources | News &
Events |
Recipes
|
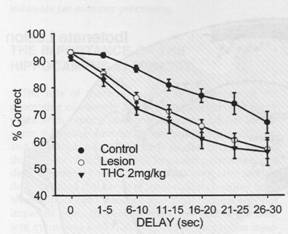 The figure to the left shows
results from a delayed
The figure to the left shows
results from a delayed 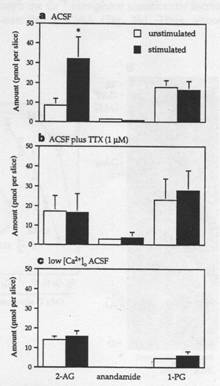 Evidence for this hypothesis begins by identifying two
Evidence for this hypothesis begins by identifying two 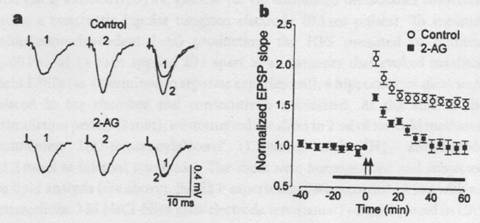
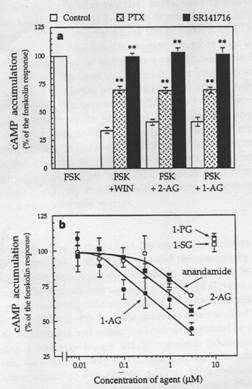 The figure to the right (a) shows the results from an experiment on cyclic AMP
levels in neurons with several different
The figure to the right (a) shows the results from an experiment on cyclic AMP
levels in neurons with several different 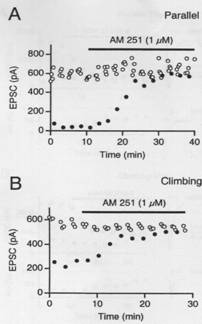
 leads to a suppression of the
leads to a suppression of the 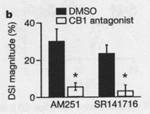 An opposite effect, depolarization-induced suppression of inhibition (DSI) has
been shown to occur in
An opposite effect, depolarization-induced suppression of inhibition (DSI) has
been shown to occur in